However, at day 14 of the estrous cycle only subtle differences were observed in their transcriptomes by endometrial receptivity. To refine these results, and to obtain information about expression earlier in estrous, we performed endometrial expression profiling at days 3 and 7 of estrous in cows from high and low receptivity groups. Additionally, transcription factor analyses were conducted to identify the cascade of upstream transcriptional regulators Lomitapide Mesylate inducing gene expression changes in the endometria. Embryo implantation is a complex process. Successful implantation requires appropriate communication between the embryo and the maternal endometrium, a tissue that is crucial for the establishment and maintenance of pregnancy. A receptive endometrium and a normal and functional embryo at the blastocyst developmental stage are prerequisites of a successful implantation, which is characterized by a synchronized dialogue between maternal and embryonic tissues. However, the regulation of implantation remains incompletely understood. On one hand, the process of implantation is driven by embryonic signals conveyed through various proteins, cytokines, and growth factors that systematically modulate maternal anatomy, endocrinology, immunology, and physiology to create an environment conductive to fetal development and survival. On the other hand, however, inadequate uterine receptivity is thought to be responsible for two-thirds of implantation failure. A recent report described transcriptional changes in the endometrium during the menstrual cycle and embryonic implantation in human. Similarly, expression profiling has been performed during both the estrous cycle and implantation in cattle, even Gomisin-D accounting for different biotechniques used to derive embryos. These reports identified the endometrium as an early sensor for the presence and implantation of embryos in the uterus. The microarray data reported here indicate that endometrial expression patterns and alterations of transcript abundance are indicative of the receptivity of the endometrium and, thus, of pregnancy success. Several molecules involved in integrin signaling, cytoskeleton signalling, and Rac signaling were enriched in the high receptive endometrium at the beginning of the estrous cycle. Transcripts of these pathways were found to increase during the window of implantation and were proposed as markers for the implantation process. Integrins participate in cell�Cmatrix and cell�Ccell adhesion in many physiologically important processes. Cellular aggregation leads to the recruitment of a network of cytoskeletal proteins and intracellular signaling complexes. The acquisition of adhesion ligands, together with the loss of inhibitory components that may act as a barrier to an attaching embryo, contribute to endometrial receptivity. In cattle a number of mRNAs coding for components of the cytoskeleton and extracellular matrix proteins were as enriched at estrous. Our study shows that genes of molecular signaling pathways that determine the communication skills of the endometrium are differentially expressed between the high and low receptive groups at a time early in advance of implantation. Many transcripts showed lower abundance in the low receptive endometrium at 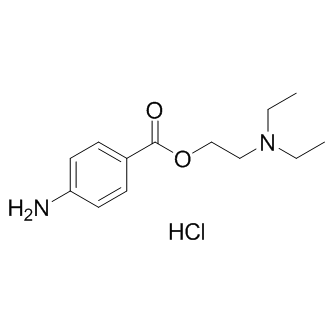 day 3 of estrous belong to biofunctions relevant to energy supply, including mitochondrial pathways. This indicates that defects in these pathways may contribute to the development of low receptive endometrium. The role of estrogen in directly and indirectly affecting mitochondrial activity is well established and was functionally exploited in experiments using estrogen receptora knockout mice and ovariectomized rats in combination with estrogen supplementation. Others reported that estrogen increases NRF-1 transcription, resulting in a coordinated increase of expression of nuclear and mitochondrial-encoded genes.
day 3 of estrous belong to biofunctions relevant to energy supply, including mitochondrial pathways. This indicates that defects in these pathways may contribute to the development of low receptive endometrium. The role of estrogen in directly and indirectly affecting mitochondrial activity is well established and was functionally exploited in experiments using estrogen receptora knockout mice and ovariectomized rats in combination with estrogen supplementation. Others reported that estrogen increases NRF-1 transcription, resulting in a coordinated increase of expression of nuclear and mitochondrial-encoded genes.
Author: screening library
Interventions designed for younger subjects may have off-target effects in aged subjects
Sox9 controls the determination of cartilage and muscle cell fate in muscle satellite cells, and may play important roles in regulating the process of fracture healing. As these factors are also involved in early embryonic cell fate determination, our work supports the notion that healing 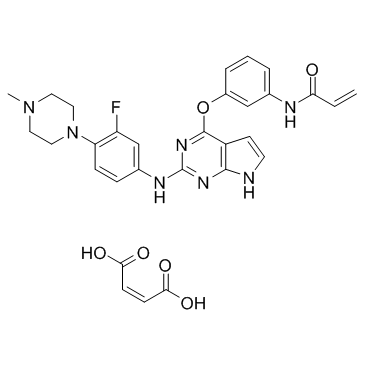 recapitulates development. Understanding these signaling events may eventually allow us to harness these mechanisms of chondrogenic differentiation to enhance fracture healing. Age-related cognitive deficits are highly prevalent and constitute an important health risk in the human population. They can presage development of neurodegenerative disease, and are a primary reason for elderly placement in assisted living facilities. Perturbations in sleep are also a common complaint among the elderly and include circadian advance, sleep fragmentation, and insomnia. Healthy young adults show some aging-like phenotypes when deprived of sleep, including daytime sleepiness, Chlorhexidine hydrochloride metabolic syndrome-like changes, and cognitive deficits. This is consistent with work suggesting that sleep promotes memory, possibly through slow wave influence on synapses and/or promotion of macro molecule synthesis. Further, numerous studies have pointed to the deleterious effects of stress and stress hormones on brain function and a major hypothesis of aging posits that continued exposure to stress and stress hormones over age is a fundamental cause of age-related deficits in various systems. Thus, dysregulated sleep and stress seen with age might contribute to age-related functional changes. Despite the seemingly similar effects of age, stress and sleep deprivation, and the high prevalence of sleep changes and new onset stress with age, relatively few studies have tested for a molecular relationship between the influences of SD, stress, and aging on brain tissue. Here, we hypothesized that an aged or stressed animal’s hippocampal transcriptional profile would be similar to that of a sleep-deprived subject. We tested a prediction of this hypothesis by sleep depriving young animals and statistically testing for aging- or stress-like transcriptional phenotypes in the hippocampus. Young F344 rats were sleep deprived for 24 or 72 hours using the modified multi-platform ‘flower pot’ method. Blood corticosterone levels, adrenal weights, body weights, and hippocampal CA1 gene expression profiles were measured. A second set of rats was exposed to novel environment stress for 24 or 72 hours to help control for non-specific stress effects of the environmental change necessitated by the sleep deprivation protocol. In a third set of animals, the SD protocol was applied and gene products were validated at the protein level using Western blots. Data were subjected to bioinformatic analysis and contrasted with results from prior transcriptional profiling studies as noted in Results. Transcriptional comparison suggests SD, stress and aging interact with a similar subset of genes within the hippocampal transcriptome. However, although there was strong directional agreement between SD and NES, notable disagreements between SD and aging were seen, including opposite inflammatory and glial expression changes. Our studies also identified SD-specific genes and gene profiles that may represent targets for therapeutic intervention. These include the previously identified Homer1 as a potential sleep regulation molecule, but also other novel Tulathromycin B candidate genes involved in pathways related to synaptic function, vesicular release and intercellular adhesion.
recapitulates development. Understanding these signaling events may eventually allow us to harness these mechanisms of chondrogenic differentiation to enhance fracture healing. Age-related cognitive deficits are highly prevalent and constitute an important health risk in the human population. They can presage development of neurodegenerative disease, and are a primary reason for elderly placement in assisted living facilities. Perturbations in sleep are also a common complaint among the elderly and include circadian advance, sleep fragmentation, and insomnia. Healthy young adults show some aging-like phenotypes when deprived of sleep, including daytime sleepiness, Chlorhexidine hydrochloride metabolic syndrome-like changes, and cognitive deficits. This is consistent with work suggesting that sleep promotes memory, possibly through slow wave influence on synapses and/or promotion of macro molecule synthesis. Further, numerous studies have pointed to the deleterious effects of stress and stress hormones on brain function and a major hypothesis of aging posits that continued exposure to stress and stress hormones over age is a fundamental cause of age-related deficits in various systems. Thus, dysregulated sleep and stress seen with age might contribute to age-related functional changes. Despite the seemingly similar effects of age, stress and sleep deprivation, and the high prevalence of sleep changes and new onset stress with age, relatively few studies have tested for a molecular relationship between the influences of SD, stress, and aging on brain tissue. Here, we hypothesized that an aged or stressed animal’s hippocampal transcriptional profile would be similar to that of a sleep-deprived subject. We tested a prediction of this hypothesis by sleep depriving young animals and statistically testing for aging- or stress-like transcriptional phenotypes in the hippocampus. Young F344 rats were sleep deprived for 24 or 72 hours using the modified multi-platform ‘flower pot’ method. Blood corticosterone levels, adrenal weights, body weights, and hippocampal CA1 gene expression profiles were measured. A second set of rats was exposed to novel environment stress for 24 or 72 hours to help control for non-specific stress effects of the environmental change necessitated by the sleep deprivation protocol. In a third set of animals, the SD protocol was applied and gene products were validated at the protein level using Western blots. Data were subjected to bioinformatic analysis and contrasted with results from prior transcriptional profiling studies as noted in Results. Transcriptional comparison suggests SD, stress and aging interact with a similar subset of genes within the hippocampal transcriptome. However, although there was strong directional agreement between SD and NES, notable disagreements between SD and aging were seen, including opposite inflammatory and glial expression changes. Our studies also identified SD-specific genes and gene profiles that may represent targets for therapeutic intervention. These include the previously identified Homer1 as a potential sleep regulation molecule, but also other novel Tulathromycin B candidate genes involved in pathways related to synaptic function, vesicular release and intercellular adhesion.
A chemopreventive effect of Resv whereas increased breast cancer incidence is observed in one study
The reason for these contradictory results is not obvious. Resveratrol reduced chemically-induced liver cancer in rats and in mice 4-(Benzyloxy)phenol injected with carcinoma cells. A single Catharanthine sulfate experiment showed no reduction of the incidence of esophagus carcinoma formation by Resv. The effect of Resv on gastric cancers was evaluated with its effect on tumor growth after subcutaneous injection of cancer cells. Both studies in mice showed a reduced tumor volume as a consequence of Resv exposure. To study the effect of Resv on colon cancer development in animals, four studies focus on chemically-induced colon cancer in rats or mice and three studies evaluated the effect of Resv on colon cancer in APC/Min mice strains. Three of the tests showed significant reduction of the aberrant crypt formation or incidence of adenomas, whereas two of the mouse studies did not show a significant reduction in colon cancer incidence or tumor load. Resveratrol showed a chemopreventive effect on development of prostate cancer using rat or mice strains prone to spontaneously developing prostate cancer and a mouse model with injected prostate cancer cells In other studies focusing on lung carcinogenesis, Resv showed a chemopreventive effect only in a single experiment out of a total of four experiments. A single study 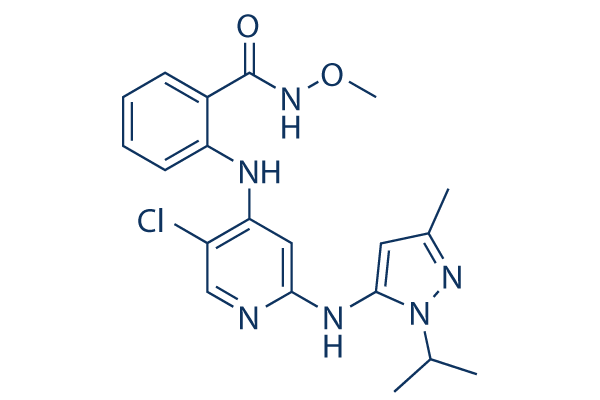 focused on the effect of Resv on the development of neuroblastoma in mice, and found reduced tumor volume. Several animal studies have indicated that Resv has a neuroprotective effect. In total, 12 studies test the effect of resveratrol after a single exposure and 28 studies investigated the effect of resveratrol af subchronic/chronic exposure. This effect of Resv has been documented in various animal models including rabbits, mice or rats and using different end points, such as reduced lipid peroxidation and neurological cell destruction, attenuation of induced lesion areas, induced tolerance to brain injury, reduced frequency of seizures, impairment of motor coordination and enhancement of learning. Only very few of these experiments found no or a marginal effect of Resv. Inflammatory response is a well known mechanism of the diseases described above such as cancer, coronary-heart disease, diabetes and neurodegeneration. Resveratrol is shown to modulate the inflammatory response induced by various stimuli. Fourteen studies have investigated the effect of exposure of one week or more to Resv on various inflammatory markers in rats. The same number of studies has been identified using mice as an experimental model. Generally, Resv in nearly all models counteracted the increased levels of pro inflammatory biochemical markers, such as TNFa, IL-1b, IL-6 in nearly all models. Beside these cytokines, MCP-1, COX-2 and iNOS was most often found to be downregulated by Resv when stimulated by the pro-inflammatory treatment. The estimates of inflammation were often performed as a part of a study with another aim, i.e. testing a chemopreventive effect, or the effect of Resv on diabetes or cardiovascular disease. Therefore, different inducers of the inflammatory status have been used; Resv reduced inflammation in several models such as obesityinduced, in diabetic mice or chemically induced diabetes, but also dextran sulfate sodium induced colon colitis. Other models were using induced hypertension, chemicals causing tissue injury in liver, lung and colon and showed decreased levels of inflammatory markers. Besides the animal experiments described above which focused on chronic or near-chronic exposures, a long list of papers exist that analyze the effect of Resv after an acute exposure on biomarkers relevant for prevention.
focused on the effect of Resv on the development of neuroblastoma in mice, and found reduced tumor volume. Several animal studies have indicated that Resv has a neuroprotective effect. In total, 12 studies test the effect of resveratrol after a single exposure and 28 studies investigated the effect of resveratrol af subchronic/chronic exposure. This effect of Resv has been documented in various animal models including rabbits, mice or rats and using different end points, such as reduced lipid peroxidation and neurological cell destruction, attenuation of induced lesion areas, induced tolerance to brain injury, reduced frequency of seizures, impairment of motor coordination and enhancement of learning. Only very few of these experiments found no or a marginal effect of Resv. Inflammatory response is a well known mechanism of the diseases described above such as cancer, coronary-heart disease, diabetes and neurodegeneration. Resveratrol is shown to modulate the inflammatory response induced by various stimuli. Fourteen studies have investigated the effect of exposure of one week or more to Resv on various inflammatory markers in rats. The same number of studies has been identified using mice as an experimental model. Generally, Resv in nearly all models counteracted the increased levels of pro inflammatory biochemical markers, such as TNFa, IL-1b, IL-6 in nearly all models. Beside these cytokines, MCP-1, COX-2 and iNOS was most often found to be downregulated by Resv when stimulated by the pro-inflammatory treatment. The estimates of inflammation were often performed as a part of a study with another aim, i.e. testing a chemopreventive effect, or the effect of Resv on diabetes or cardiovascular disease. Therefore, different inducers of the inflammatory status have been used; Resv reduced inflammation in several models such as obesityinduced, in diabetic mice or chemically induced diabetes, but also dextran sulfate sodium induced colon colitis. Other models were using induced hypertension, chemicals causing tissue injury in liver, lung and colon and showed decreased levels of inflammatory markers. Besides the animal experiments described above which focused on chronic or near-chronic exposures, a long list of papers exist that analyze the effect of Resv after an acute exposure on biomarkers relevant for prevention.
The molecular details of receptor activation limitation include receptor misfolding and constraining of the receptor into conformational
In contrast, antibodies, both polyclonal and monoclonal, have exquisite conformational specificity and extensively used in hormone-receptor interactions studies. Our earlier studies using agonistic FSHR hinge antibody as well as Cinoxacin Extensive use of polyclonal and monoclonal antibodies in investigation of structure-function relationship of TSHR and its ligands clearly indicate the expediency of using such antibodies in studying the role of hinge region in receptor activation. In the present study, we have combined mutagenesis and novel TSHR MAbs to dissect out roles of different regions of the receptor in binding and signaling. Effect of MAbs on hTSH-TSHR interactions was further investigated by determining binding constants in presence of MAbs. Typical TSH radioreceptor assays were carried out after pre-incubating the receptor preparation with MAbs or NMIgG and the binding data was converted to the Scatchard plots and the binding constants calculated according to the legends of Figure 3. The Scatchard analysis of TSH-TSHR binding is typically curvilinear and for visual representation, only the physiologically relevant high affinity receptor component has been considered. The effect of the antibodies on the low affinity, high capacity components is, however, presented in Figure S10. As shown in Figure 3 and Table 1, MAbs 311.82 and 311.174 did not affect the TSH-receptor interactions, although a marginal decrease in the Bmax was observed in case of the latter MAb. The MAb 311.87 exhibited non-competitive inhibition with a significant decrease in Bmax and no change in the affinity. Extensive mutational analysis of the hinge region residues has shown the importance of the hinge region in hormone binding and receptor stimulation. These mutations, while providing valuable insights into receptor functioning, suffer from the stabilization of a Hammond state in the structural segment containing the mutation. By increasing or decreasing the energy barrier of activation, the mutation could allosterically stabilize a receptor conformation unsuitable for hormone binding, which may be misconstrued to be a part of the hormone-binding site. 3,4,5-Trimethoxyphenylacetic acid Moreover, substitution of residues at the hinge region and LRR domain junction by charged residues cause misfolding of TSHR causing impairment of cell surface expression. MAbs on the other hand have the unique capability of differentiating between an allosteric and an orthosteric receptor site and have proved to be reliable tools in detecting the ligand induced conformational changes in the wild type or mutated receptors. TSHR stimulatory monoclonal antibodies have been instrumental in understanding TSH interaction with the LRR domain of TSHR or the N terminal region of ECD in receptor activation. Almost all the reported TSH stimulatory MAbs had epitopes in the LRR domain except TSAB-4 that recognized in the TSHR hinge region or all were full agonists except IRI-SAb1 and affected TSH binding. In contrast, MAb 311.62 is a hinge region specific partial agonistic MAb that does not inhibit TSH binding but stimulates the receptor. In the present study, MAB 311.62 and the antagonistic MAB 311.87 have been used to understand the mechanisms of hormone binding and receptor activation. It is expected that transmission of signal from the 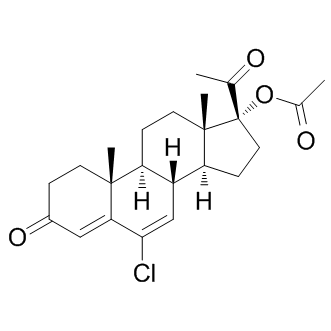 LRR to the TMD would require considerable conformational change in the receptor. It is highly unlikely that contact of the hormone at LRR 4�C6 alone can initiate such a conformational change owing to its rigid structure.
LRR to the TMD would require considerable conformational change in the receptor. It is highly unlikely that contact of the hormone at LRR 4�C6 alone can initiate such a conformational change owing to its rigid structure.
Though gain mutations in GpHRs has been a popular approach to by current wake-promoting agents
Seems a valid target for the development of future sleep countermeasures. No currently available sleep countermeasures allow human ‘normal wake’ performance over extended periods of time. Frank limitations include dissociative/psychotic behavior after prolonged exposure, risk of developing tolerance/dependence on the agents being used, and lowered seizure thresholds. Our microarray results indicate that synaptic function-related mRNA expression changes do not occur in a vacuum. Among other categories of change, the reduction in intercellular adhesion molecule expression may be of particular importance. These molecules are pleiotropic and in addition to playing a role in inflammation and immune signaling, also help maintain synaptic juxtaposition. We speculate that their downregulation in SD could leading to aberrant synaptic signaling. Thus, the development of drugs targeting the stabilization of intercellular adhesion molecules may help to extend the duration of action for stimulant-based sleep countering agents.1. Our findings support long-standing observations that SD elicits physiologic and transcriptional responses with stress-like features. Neuronal synaptic gene expression changes may help to explain the benefits and limitations of current stimulant-based SD-countering therapeutics and points to novel targets in presynaptic vesicular release and neural cell adhesion for development of future SD-countering drugs. Significantly more genes than expected by chance were commonly regulated by both aging and SD. Although many were driven in the same direction by these two phenomena, we could not fully support the hypothesis that SD evokes an aging-like transcriptional change. Genes showing significant but opposite expression changes in SD and aging, including Chrbp, Fkbp4, Homer1 and Per2, are candidates for further study of the interplay between SD and aging. Compared to prior studies showing increased inflammatory signaling with stress, our novel environment stress protocol did not influence inflammatory gene expression to an appreciable extent, and SD suppressed those signals. This suggests that SD in our hands was either a weak stressor, or that it has an effect mechanistically distinct from that of stress on immune signaling. One of the receptor activation models suggests that the C-terminal region of ECD, which links LRRs to TMD, acts as a flexible “hinge”, enabling the hormone bound LRR to communicate with the Extracellular loops of TMD. The scaffold model, however, does not explain the cryptic hormone binding sites in the hinge region of FSHR or the interaction of the a-subunit of hormones with the hinge regions. Removal of ECD by trypsin treatment, or mutations or deletions in the hinge region cause an increase in hormone-independent activation by releasing the silencing effect of the ECD and have since given rise to the tethered inverse agonist model in which ECD Gomisin-D serves as an inverse agonist stabilizing TMD in an inactive conformation. Hormone binding or activating mutations in the hinge region disengages these inhibitory ECD-TMD Lomitapide Mesylate interactions. However, this model is inadequate to explain relatively low activation of hinge region constitutively activating mutations like K287A or K291A compared to that  caused by the hormone even with ample expression of these mutants on cell surface. It also does not explain lack of hormone-independent activation of LHR or why S277Q in LHR responds to hCG but TSHR homologous mutation S281Q does not respond to TSH, although certain mutations in the critical S281 residue has the potential of stimulating TSHR comparable to the hormone.
caused by the hormone even with ample expression of these mutants on cell surface. It also does not explain lack of hormone-independent activation of LHR or why S277Q in LHR responds to hCG but TSHR homologous mutation S281Q does not respond to TSH, although certain mutations in the critical S281 residue has the potential of stimulating TSHR comparable to the hormone.