Again, as noted above for ACTH treatment, the expression levels of miRNAs differed significantly between measurements made by microarray analysis and qRT-PCR. Furthermore, a comparison of ACTH data with that of 17a-E2 data demonstrated that only,25% of the transcripts overlap. This suggests that 17a-E2induced hypocholesterolemia or direct Albaspidin-AA estrogen effects on the adrenal, but not increased ACTH secretion, is most likely responsible for the Folinic acid calcium salt pentahydrate observed alterations in the levels of specific miRNAs in adrenals of 17a-E2-treated rats. The hypothalamus-pituitary-adrenal axis consists of a set of direct influences and feedback responses between the hypothalamus, the pituitary gland and the adrenal that control reactions to stress and glucocorticoid secretion. Glucocorticoid secretion by the adrenal cortex inhibits the functions of both the hypothalamus and the pituitary gland by a negative feedback mechanism. This reduces the secretion of CRH and vasopressin and directly reduces the cleavage of pro-opiomelanocortin into ACTH and ?endorphin. In our study, we examined the impact of a synthetic glucocorticoid, dexamethasone -mediated inhibition of the HPA axis and ACTH secretion, on miRNA expression profiles in the adrenals. DEX treatment up-regulated the expression of miRNA-483, miRNA-181a-1, miRNA-490 and miRNA-181b-1, while it down-regulated the levels of miR-122, miR-466b, miR200b, miR-877, miR-296, miRNA-27a and precursor of miR-504. Furthermore, such DEX alteration of adrenal miRNA levels demonstrates that DEX suppression of endogenous ACTH secretion modulates a set of adrenal miRNAs, with the exception of miRNA-96, miRNA-466, and miRNA-27a, that are distinct from those modulated by treatment with exogenous ACTH. Interestingly, the expression of miRNA-96 is up-regulated in response to ACTH treatment, but is down-regulated following DEX treatment. Considering the current view that miRNAs act as negative regulators of gene expression, their altered expression in response to DEX may enhance and/or reduce the expression of target steroidogenic genes, leading to possibly down-regulation of adrenal steroid hormone synthesis and secretion. Our data further demonstrate that expression levels of some miRNAs are regulated by more than one hormone, i.e., by ACTH/17a-E2, ACTH/DEX, 17a-E2/DEX or 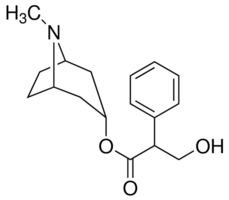 ACTH/17aE2/DEX; Table 1. The most striking similarity was observed between ACTH and 17a-E2. Both ACTH and 17a-E2 upregulated the expression of miRNA-212, miRNA-132, miRNA154, miRNA-494, miRNA-872, miRNA-194, and miRNA-24-1, but reduced the expression of miRNA-322, miRNA-20b, miRNA339, miRNA-27a, miRNA-551b, and miRNA-1224. We also observed that miRNA-30a was up-regulated in adrenals treated with ACTH, but down-regulated by 17a-E2 exposure. A comparison of effects of ACTH and DEX shows that both hormones increased the expression miRNA-181b, miRNA-672, and miRNA-100, and significantly decreased the levels of miRNA92a, and miRNA-466b. In addition to ACTH/17a-E2 and ACTH/DEX, we observed that a total of 11 miRNAs are regulated by both 17a-E2 and DEX. Among these, three mRNAs were up-regulated in response to in vivo treatment of adrenals with 17a-E2 or DEX, and the remaining eight miRNAs were downregulated in treated adrenals with either of the two hormones. Finally the expression levels of miRNA-27a and miRNA-551b were significantly reduced in adrenals of ACTH, 17a-E2 or DEX treated animals. Together, these data raise the possibility that some of these miRNAs may be intimately involved in the complex regulation of adrenal steroidogenesis.
ACTH/17aE2/DEX; Table 1. The most striking similarity was observed between ACTH and 17a-E2. Both ACTH and 17a-E2 upregulated the expression of miRNA-212, miRNA-132, miRNA154, miRNA-494, miRNA-872, miRNA-194, and miRNA-24-1, but reduced the expression of miRNA-322, miRNA-20b, miRNA339, miRNA-27a, miRNA-551b, and miRNA-1224. We also observed that miRNA-30a was up-regulated in adrenals treated with ACTH, but down-regulated by 17a-E2 exposure. A comparison of effects of ACTH and DEX shows that both hormones increased the expression miRNA-181b, miRNA-672, and miRNA-100, and significantly decreased the levels of miRNA92a, and miRNA-466b. In addition to ACTH/17a-E2 and ACTH/DEX, we observed that a total of 11 miRNAs are regulated by both 17a-E2 and DEX. Among these, three mRNAs were up-regulated in response to in vivo treatment of adrenals with 17a-E2 or DEX, and the remaining eight miRNAs were downregulated in treated adrenals with either of the two hormones. Finally the expression levels of miRNA-27a and miRNA-551b were significantly reduced in adrenals of ACTH, 17a-E2 or DEX treated animals. Together, these data raise the possibility that some of these miRNAs may be intimately involved in the complex regulation of adrenal steroidogenesis.