The data also show that NANOG dimerization plays a role: when this property is removed from the full model the immediate effect is a loss of bistability. Finally, cooperative binding of OCT4 and SOX2 in the models does not play a significant role: its loss from row 1 to row 5 caused no loss of bistability. It is also interesting to note that epigenetic coperativity alone can be a mechanism for bistability, without requiring dimerization cooperativity or cooperative binding of transcription factors. The data indicate that mechanisms can act together to bring about bistability and multiple contributing mechanisms may lead to greater robustness of bistability across varying parameters. The above results were all obtained with the deterministic ODE version of each model. An important goal of the current study is to use 20S-Notoginsenoside-R2 Stochastic simulation methodology to explore cell-to-cell variability in reprogramming dynamics, as such variability is a feature of experimentally observed reprogramming protocols. Deterministic simulations produce the same behavior when started with the same initial conditions and once they reach a steady state they remain there if unperturbed. Stochastic simulations use random event generation to simulate the non-deterministic characteristics of the timing of chemical reaction events. Thus, in a stochastic framework, which can be a more realistic treatment of biochemical systems, bistability can lie along a continuum from transient to persistent. For the purpose of iPSC generation protocols, persistent bistability that switches state only when stimulated by induction protocols would be preferable. Here we have made stochastic simulations and compared bistability properties to those in the ODE framework. We simulated stochastic dynamics for the Independent Equiprobable model with the Gillespie Stochastic Simulation algorithm for initial conditions corresponding to the uninduced state, and for initial conditions corresponding to the induced state. The distribution of outcomes for each of the two initial conditions is shown in Figure 3C and D. As expected, the data show that the model has two different distributions that depend on the initial conditions, with simulations beginning near the induced or uninduced state generally remaining there. Next, we investigated whether the bistable system could be induced from the low steady state to the high steady state by adding an exogenous source of SOX2 and OCT4. Simulations were carried out with mRNA for these genes produced at a constant rate to emulate viral- or plasmid-based induction, as is done experimentally. The results of hundreds of stochastic 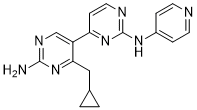 simulations demonstrate all simulations can eventually reach the induced state and remain there, although there was great variance in the time required for induction, ranging from about one day to several weeks of simulated time. The trajectories for two stochastic induction simulations are given in Figure 4. At t~0 the induction protocol began. In one of the illustrated simulations, the system changes state at day one, and in the other the change happens at day six. It is important to note that at day 20, for both cases, the external induction is removed, but the system remains in the high state. Thus, the simulation results display marked cell-to-cell variability. Beyond the small difference expected when stochastic dynamics are simulated, dramatic variation in induction times were observed, with some cells reprogramming in about a day and others taking several weeks, although each cell was modeled Benzethonium Chloride identically.
simulations demonstrate all simulations can eventually reach the induced state and remain there, although there was great variance in the time required for induction, ranging from about one day to several weeks of simulated time. The trajectories for two stochastic induction simulations are given in Figure 4. At t~0 the induction protocol began. In one of the illustrated simulations, the system changes state at day one, and in the other the change happens at day six. It is important to note that at day 20, for both cases, the external induction is removed, but the system remains in the high state. Thus, the simulation results display marked cell-to-cell variability. Beyond the small difference expected when stochastic dynamics are simulated, dramatic variation in induction times were observed, with some cells reprogramming in about a day and others taking several weeks, although each cell was modeled Benzethonium Chloride identically.