These include K161, which forms a Schiff base with the first substrate to bind the enzyme, and a catalytic triad comprised of Y107, T44 and Y133, which are strongly conserved in all DHDPS enzymes characterized to date including S. pneumoniae. Given the clinical importance of S. pneumoniae and the rise in multi-drug resistance in this Gram-positive pathogen, the aims of this study were to Rapamycin mTOR inhibitor determine the phenotype of a DHDPS gene knock mutant of S., pneumoniae; characterize the solution properties, stability and catalytic activity of SpDHDPS; and determine the high-resolution crystal  structure of the enzyme to afford structure-based drug design strategies in future studies. Solution studies also showed Sp-DHDPS possesses significantly greater thermostability compared to Ec-DHDPS. We subsequently demonstrated using analytical ultracentrifugation that the enhanced thermostability of SpDHDPS is contributed by a 45-fold tighter tetramer-dimer dissociation constant. We therefore determined the three-dimensional structure of Sp-DHDPS using X-ray crystallography to provide insight into the enhanced thermal and thermodynamic stability. The origin of the enhanced thermal stability is clearly revealed in the resolution crystal structure of Sp-DHDPS that shows a tetrameric structure of the enzyme, consistent with our solution studies. The tertiary and quaternary structure architecture of Sp-DHDPS is very similar to Ec-DHDPS with an overall RMSD for superposition of the tetramers of 1.1 ?, as well as to other structurally characterized bacterial DHDPS enzymes;. However, significant structural differences are observed between Sp-DHDPS and Ec-DHDPS at the subunit interfaces. We show that there is an increase of 60 ?2 and 300 ?2 in solvent-inaccessible Bortezomib surface area at the ‘tight’ dimer and ‘weak’ dimer interfaces of Sp-DHDPS, respectively. This significant increase in SISA, in combination with the greater proportion of hydrogen bonding residues at the ‘tight’ dimer interface, as well as the presence of residues participating in salt bridge interactions at the ‘weak’ dimer interface, is consistent with the 45-fold lower tetramer-dimer dissociation constant for Sp-DHDPS, and to its considerably higher thermal stability. Consistent with previous studies of other DHDPS enzymes, the residues that form interactions at the ‘weak’ dimer interface are poorly conserved in SpDHDPS, whereas strong conservation is observed at the ‘tight’ dimer interface where the active sites are located. Given that recent studies show dimeric mutants of DHDPS have significantly attenuated catalytic function compared to the wild-type tetramers, the poor conservation at the ‘weak’ dimer interface offers potential for the design of pathogen-specific antimicrobial agents, particularly given that protein-protein interfaces represent highly specific drug targets. Indeed, with the increase in drug-resistant bacteria linked to the overuse and misuse of broad spectrum antibiotics, exploiting the ‘weak’ dimer interface of SpDHDPS may provide a means to negate the incidence of broad spectrum drug resistance. In conclusion, through gene knock-out studies, circular dichroism spectroscopy, analytical ultracentrifugation, dynamic light scattering, enzyme kinetics and X-ray crystallography studies, we demonstrate that Sp-DHDPS is an essential, active and thermostable tetramer. Our work offers insight into rational drug design strategies targeting multiple sites of the enzyme to afford the discovery of novel antibiotic agents with potential to negate drug resistance. Ecosystems are characterized and understood from the fundamental relationships between predator and prey. When linked vertically and horizontally these relationships form food webs, which depict how energy flows through ecosystems and demonstrate how various components of the web interact.
structure of the enzyme to afford structure-based drug design strategies in future studies. Solution studies also showed Sp-DHDPS possesses significantly greater thermostability compared to Ec-DHDPS. We subsequently demonstrated using analytical ultracentrifugation that the enhanced thermostability of SpDHDPS is contributed by a 45-fold tighter tetramer-dimer dissociation constant. We therefore determined the three-dimensional structure of Sp-DHDPS using X-ray crystallography to provide insight into the enhanced thermal and thermodynamic stability. The origin of the enhanced thermal stability is clearly revealed in the resolution crystal structure of Sp-DHDPS that shows a tetrameric structure of the enzyme, consistent with our solution studies. The tertiary and quaternary structure architecture of Sp-DHDPS is very similar to Ec-DHDPS with an overall RMSD for superposition of the tetramers of 1.1 ?, as well as to other structurally characterized bacterial DHDPS enzymes;. However, significant structural differences are observed between Sp-DHDPS and Ec-DHDPS at the subunit interfaces. We show that there is an increase of 60 ?2 and 300 ?2 in solvent-inaccessible Bortezomib surface area at the ‘tight’ dimer and ‘weak’ dimer interfaces of Sp-DHDPS, respectively. This significant increase in SISA, in combination with the greater proportion of hydrogen bonding residues at the ‘tight’ dimer interface, as well as the presence of residues participating in salt bridge interactions at the ‘weak’ dimer interface, is consistent with the 45-fold lower tetramer-dimer dissociation constant for Sp-DHDPS, and to its considerably higher thermal stability. Consistent with previous studies of other DHDPS enzymes, the residues that form interactions at the ‘weak’ dimer interface are poorly conserved in SpDHDPS, whereas strong conservation is observed at the ‘tight’ dimer interface where the active sites are located. Given that recent studies show dimeric mutants of DHDPS have significantly attenuated catalytic function compared to the wild-type tetramers, the poor conservation at the ‘weak’ dimer interface offers potential for the design of pathogen-specific antimicrobial agents, particularly given that protein-protein interfaces represent highly specific drug targets. Indeed, with the increase in drug-resistant bacteria linked to the overuse and misuse of broad spectrum antibiotics, exploiting the ‘weak’ dimer interface of SpDHDPS may provide a means to negate the incidence of broad spectrum drug resistance. In conclusion, through gene knock-out studies, circular dichroism spectroscopy, analytical ultracentrifugation, dynamic light scattering, enzyme kinetics and X-ray crystallography studies, we demonstrate that Sp-DHDPS is an essential, active and thermostable tetramer. Our work offers insight into rational drug design strategies targeting multiple sites of the enzyme to afford the discovery of novel antibiotic agents with potential to negate drug resistance. Ecosystems are characterized and understood from the fundamental relationships between predator and prey. When linked vertically and horizontally these relationships form food webs, which depict how energy flows through ecosystems and demonstrate how various components of the web interact.
Author: screening library
Growth in a high-temperature environment and likely modulates the SakA MAPK phosphorylation
Taken together with findings in this study, it appears that a robust and complex signal transduction system for adaptation to extracellular environments exists in A. fumigatus. In conclusion, we found that NikA plays an important role in conidiation, morphology, and stress responses, and that the SakA MAPK cascade is regulated through SskA in response to osmotic shock and fungicide treatment. We also characterized the other HKs including Fos1, PhkA, PhkB, Fhk5, and Fhk6. There were no interactions between these HKs and NikA or SakA, at least under the conditions tested here. Although NikA seems to be dispensable in A. fumigatus pathogenicity, molecular insights provided in this study may open possibilities in the NVP-BKM120 944396-07-0 development of new antifungal chemicals focusing on the TCS signaling of medically relevant fungal pathogens. Bacterial pathogens subvert host eukaryotic cellular pathways for survival and replication; in turn, infected host cells respond to the invading pathogen through cascading changes in gene expression. Deciphering these complex temporal and spatial dynamics to identify novel bacterial virulence factors or host response pathways is crucial for improved diagnostics and therapeutics. Microarrays have been the predominant methodology for determining gene expression profiles, revealing a diversity of bacterial pathogenic mechanisms and commonalities of the complex global host response to infection. However, microarrays are inadequate for profiling both prokaryotic and eukaryotic RNA from infected cells, as they are limited to what can be printed and detected on the array. Technical limitations such as high background signals and cross-hybridization also limits their dynamic range. Consequently, array analyses of hostpathogen interactions have typically examined either the pathogen or the host, but usually not both simultaneously. The few studies that examine both bacterial and host cell transcriptional responses separate the prokaryotic and eukaryotic messenger RNA prior to microarray profiling. Sufficient prokaryotic mRNA for hybridization can be difficult to obtain unless axenic culture or selective amplification is used or, in the case of intracellular bacteria, in vitro infections are established with high multiplicities of infection. High MOIs may not represent natural infection levels, distorting expression profiles. The early events following invasion are often poorly characterized, as the small number of organisms yields insufficient transcripts for microarray detection. Furthermore, standard microarrays are restricted to ALK5 Inhibitor II citations existing genome annotation and cannot detect novel RNA moieties that are not printed on the array. Tiling arrays overcome this limitation and have been successfully applied to bacteria, revealing antisense RNA expression and other non-coding RNA transcripts. However, the large size of eukaryotic genomes makes tiling arrays prohibitively expensive for host gene expression studies. Tag-based sequencing methods alleviate these problems to some extent, allowing individual transcripts to be digitally counted with a broad dynamic range. Nevertheless, as these approaches only sample a small region of a transcript, they cannot capture the full diversity of RNA classes and isoforms. RNA-Seq, or deep sequencing of cDNA libraries by nextgeneration sequencing, circumvents many of the problems associated with microarray profiling or tag-based sequencing. RNA-Seq can comprehensively and systematically define the transcriptome of an organism with minimal bias, across different experimental conditions or cell types without  probe design or cross-hybridization problems. RNA-Seq data are consistent with microarray results but are more sensitive, with essentially an infinite dynamic range. RNA-Seq is annotationindependent, allowing novel transcript discovery without being reliant on array design or preexisting annotation.
probe design or cross-hybridization problems. RNA-Seq data are consistent with microarray results but are more sensitive, with essentially an infinite dynamic range. RNA-Seq is annotationindependent, allowing novel transcript discovery without being reliant on array design or preexisting annotation.
Among the paired aliquots from individual samples relatively poor pairwise correlation
Inspection of the corresponding scatter plot showed many data points above and below the primary linear cluster. Similarly, the heat map in Fig. 2B showed uniformly poor correlations for P3, and the scatter plot comparing pooled sample aliquots P3 vs. P4 displayed a group of points with higher values in the P4 sample than in P3. To investigate the source of this variability, we examined the sequences of peptides that were poorly correlated in pairwise comparisons and considered the proteins from which they were derived. Examination of the identified proteins revealed that some samples contained “process contaminant” proteins. The peptides from these proteins were removed from subsequent analyses of all samples and a new correlation heat map was generated. Although this exclusion of keratins and MAF proteins improved the correlations between selected pairwise comparisons, it did not improve all poor correlations. Further examination of the scatter plot of annotated peptide intensities for one of these unchanged pairwise comparisons showed a distinct cloud of points with lower values in  sample 1a. All the sequences of these discordant peptides contained at least one methionine residue. Removal of these methionyl peptides from subsequent PCC calculations resulted in an increased correlation coefficient for 1a vs. 1b, and a modest increase in the correlation coefficient for 5a and 5b. However, the removal of methionyl peptides did not significantly alter the poorer correlation between P3 and the other pooled samples. A scatter plot of the peptide intensities of P3 vs. P4 showed a discrete group of peptide intensities that were lower in aliquot P3 than in P4. Upon inspection, these discordant peptides were found to have early Reversine elution times during liquid chromatography. The markedly decreased ion current for P3 compared to P4 during this early elution window is shown in Fig. S4, panels A and B. As an example of the marked reduction in signal for P3 during this elution time window, all sample intensities for an early-eluting NCAM-1 peptide are shown in Fig. S4, panels C and D. Scatter plots of the peptide intensities of P3 vs. P4 before and after removal of the early eluting peptides show improved correlation after exclusion. In kind, a new non-clustering heat map of PCC values, created after excluding these early-eluting peptides from all PCC calculations, showed improved correlations and a narrower range of PCC values between P3 and all other pooled samples. Although the reason for the low intensities of earlyeluting peptides found in sample P3 remains unclear and does not appear to affect other samples substantially, these early-eluting peptides were excluded from subsequent analyses of all samples. To estimate the residual technical variability with which this technique quantifies each of these proteins, coefficients of variation were calculated from replicates of the pooled CSF sample; to determine the inter-individual variability of each of these proteins within a group of cognitively normal individuals, subject variance was calculated from the paired aliquots from individual samples. These assessments of variability are important for biomarker studies because, hypothetical proteins with relatively higher CVs and/or relatively high subject variance among controls will be more likely than those with lower CVs and lower subject variances to show overlapping ranges between diseased and control cohorts, given equivalent fold-changes associated with disease. Greater overlaps mean lower sensitivity and specificity and, therefore, relatively less potential for a candidate to serve as an effective disease biomarker. This study evaluates the technical merits and potential of a nonbiased proteomics technique, label-free MK-1775 quantitative LC-MS/MS.
sample 1a. All the sequences of these discordant peptides contained at least one methionine residue. Removal of these methionyl peptides from subsequent PCC calculations resulted in an increased correlation coefficient for 1a vs. 1b, and a modest increase in the correlation coefficient for 5a and 5b. However, the removal of methionyl peptides did not significantly alter the poorer correlation between P3 and the other pooled samples. A scatter plot of the peptide intensities of P3 vs. P4 showed a discrete group of peptide intensities that were lower in aliquot P3 than in P4. Upon inspection, these discordant peptides were found to have early Reversine elution times during liquid chromatography. The markedly decreased ion current for P3 compared to P4 during this early elution window is shown in Fig. S4, panels A and B. As an example of the marked reduction in signal for P3 during this elution time window, all sample intensities for an early-eluting NCAM-1 peptide are shown in Fig. S4, panels C and D. Scatter plots of the peptide intensities of P3 vs. P4 before and after removal of the early eluting peptides show improved correlation after exclusion. In kind, a new non-clustering heat map of PCC values, created after excluding these early-eluting peptides from all PCC calculations, showed improved correlations and a narrower range of PCC values between P3 and all other pooled samples. Although the reason for the low intensities of earlyeluting peptides found in sample P3 remains unclear and does not appear to affect other samples substantially, these early-eluting peptides were excluded from subsequent analyses of all samples. To estimate the residual technical variability with which this technique quantifies each of these proteins, coefficients of variation were calculated from replicates of the pooled CSF sample; to determine the inter-individual variability of each of these proteins within a group of cognitively normal individuals, subject variance was calculated from the paired aliquots from individual samples. These assessments of variability are important for biomarker studies because, hypothetical proteins with relatively higher CVs and/or relatively high subject variance among controls will be more likely than those with lower CVs and lower subject variances to show overlapping ranges between diseased and control cohorts, given equivalent fold-changes associated with disease. Greater overlaps mean lower sensitivity and specificity and, therefore, relatively less potential for a candidate to serve as an effective disease biomarker. This study evaluates the technical merits and potential of a nonbiased proteomics technique, label-free MK-1775 quantitative LC-MS/MS.
Follicular fluid originate from different cell types within follicular microenvironment
We have investigated the expression of 8 SCH772984 selected candidate miRNAs in granulosa cells, theca cells and cumulus oocyte complex. The results revealed that all these miRNAs are detected in all follicular cell types with varying expression level. MicroRNAs like miR-640, miR-526b* and miR-381 were abundant at higher level in theca cells, while miR-373, miR-30e* and miR-19b-1* expressed more in COCs. However, no significant differences were observed across different cell types in the expression of miR-654-5p and miR-29c. Although the origin of extra-cellular miRNAs in body fluids remains elusive, several reports demonstrated that the origin of extra-cellular miRNAs is closely related to the surrounding cells. While a common set of miRNAs were found in equine follicular fluid and surrounding follicular cells, blood cells are reported to be the major contributor of circulating miRNAs in serum. Therefore, the SAR131675 1433953-83-3 presence of significant level of candidate extra-cellular miRNAs in the surrounding follicular cells could enable us to postulate that the majority of extra-cellular miRNAs in follicular fluid originated from various cell types in the course of their communication during oocyte growth. In order to elucidate the possible exchange of molecular signals including miRNAs in form of exosome between follicular cells we have investigated the ability of granulosa cells to take up exosomes isolated from follicular fluid. Prior to uptake experiment, the stability of exosome coupled miRNA under in vitro culture environment was determined and result showed that exosome coupled miRNAs were stable not only up to 24 hours but even until 48. Similar studies have also shown the stability of circulating serum miRNAs, breast milk exosomal miRNA at room temperature and multiple freeze-thaw cycle. This may provide a great potential for functional analysis of exosome mediated transport of molecules for various cellular processes under in vitro conditions. Following the confirmation of exosomal miRNA stability under culture conditions, we performed exosomes uptake experiment by primary granulosa cells. Fresh exosomes isolated from follicular fluid and labeled by PKH67 fluorescent dye were co-cultured with bovine primary granulosa cells in vitro for 24 hrs. The fluorescent microscopy observation of labeled exosomes co-cultured granulosa cells revealed the presence of green fluorescent exosomes in cultured primary granulosa cells. Similar observation was reported in equine granulosa cells, microglial cell line, immune cells and other cell lines. Fluorescent microscopy result also shows that the majority of the exosomes taken up by granulosa cells were gathered around perinuclear region. Importantly, there were no fluorescent signals in plasma membrane indicating that exosomes were internalized in to granulosa cells via endocytosis. If fusion was the dominant pathway for exosomes uptake, plasma membrane would contain fluorescent signal after co-culture of cells and labeled exosomes. Similar results were  observed in exosomes internalization by resting PC12 cells. The consequence of transfection of exosome-coupled miRNAs in endogenous miRNA abundance in granulosa cells was investigated by expression analysis of candidate miRNAs in exosomes derived from follicular fluid of either BCB+ or BCB- oocyte origin. Results have evidenced increased level of endogenous miRNA in exosome transfected granulosa cells compared to the control ones. This could enable us to conclude that exosome mediated exchange of miRNAs in follicular microenvironment is an important way of communication between follicular cells. However, the transfer of other miRNAs through these exosomes and their enrichment in granulosa cells and their subsequent contribution to the regulation of the same target genes or other genes cannot be ruled out.
observed in exosomes internalization by resting PC12 cells. The consequence of transfection of exosome-coupled miRNAs in endogenous miRNA abundance in granulosa cells was investigated by expression analysis of candidate miRNAs in exosomes derived from follicular fluid of either BCB+ or BCB- oocyte origin. Results have evidenced increased level of endogenous miRNA in exosome transfected granulosa cells compared to the control ones. This could enable us to conclude that exosome mediated exchange of miRNAs in follicular microenvironment is an important way of communication between follicular cells. However, the transfer of other miRNAs through these exosomes and their enrichment in granulosa cells and their subsequent contribution to the regulation of the same target genes or other genes cannot be ruled out.
In addition DprB is a dehydrin-like protein responsible in morphology and pathogenicity
In addition, the afskn7 deletion mutant constructed by our group showed no significant resistance to cell wall-perturbing reagents and fungicides. Thus, to better understand how NikA regulates conidiation, morphology, and responses to chemicals, the generation and investigation of a double deletion mutant of sskA and afskn7 is necessary. This 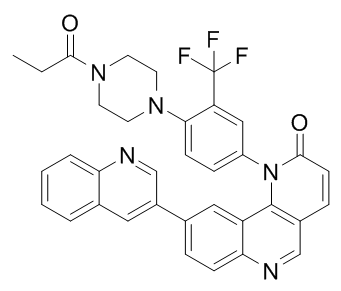 will be the subject of our future study. As in other fungi, the A. fumigatus SakA MAPK was transiently phosphorylated by osmotic shock and fungicide treatment, which was fully dependent on SskA. This clearly indicates that under the conditions tested here the SakA MAPK cascade is solely regulated by the TCS. Unexpectedly, we found that SakA was phosphorylated in response to osmotic shock in DnikA cells, suggesting that another mechanism modulates the SakA R428 in vivo phosphorylation instead of NikA, possibly via SskA. On the one hand, NikA seemed to be indispensable for the accumulation of phosphorylated SakA in response to low concentrations of fludioxonil treatment, whereas it was dispensable when treated with 10 mg/ mL fludioxonil. Judged by the roughly estimated growth ICs, 10 mg/mL of fludioxonil seems high and may cause a leaky effect at a specific interaction between the chemical and the target protein. In C. heterostrophus and N. crassa, the accumulation of phosphorylated Hog1-type MAPK was observed in the cells lacking the Nik1-type HK in the presence of high concentrations of fungicides or high osmolar reagents. These results could support the idea that another HK or mechanism contributes to the phosphorylation of SakA via SskA in some filamentous fungi. Notably, in contrast to our results, in a previous study by McCormick et al., TcsC/ NikA has been shown to be required for the phosphorylation of SakA in response to fludioxonil treatment and osmotic shock in A. fumigatus. We think that this discrepancy is derived from the differences in media, cultivation time, and treatment time. Considering these results, the regulation of the SakA MAPK cascade by NikA may be growth stage-specific. We also found that the phosphorylation level of the SakA MAPK in DnikA was moderately high in the absence of stimuli. This increased phosphorylation level is reminiscent of a role of the S. cerevisiae Sln1p in modulating the Hog1 MAPK phosphorylation, where a defect in the sln1 gene causes abnormal activation of the Hog1 MAPK and leads to growth defects. In fact, DnikA showed a slight defect in radial growth on YGMM plates and increased the phosphorylation of SakA. These findings could support the idea that similarly to Sln1p NikA is a negative regulator of the SakA MAPK, whereas it plays a positive role at least in response to fludioxonil treatment. Here, it should be noted that the expression level of catA, dprA, and dprB in DnikA in the absence of stimuli was comparable to that in WT, suggesting that the moderate accumulation of phosphorylated SakA dose not necessarily lead to the high expression of the genes. There may be unknown regulatory mechanisms, for example a negative feedback system, concerning this. In any event, judged by the results of SakA phosphorylation and transcriptional responses to the stimuli, the phosphorylated form of SakA appears to be an active form. Also, we showed a straightforward signaling from SskA to the SakA MAPK in the A. fumigatus HOG pathway both at the transcriptional and post-translational levels. In KRX-0401 hyperosmotic stress conditions, the DnikA, DsskA, and DsakA strains showed growth sensitivity at distinct levels. As has been well studied in S. cerevisiae, the HOG pathway plays an important role in osmotic adaptation by regulating glycerol accumulation; thus, it is possible that glycerol accumulation occurred in response to hyperosmotic stress conditions in A. fumigatus, and the HOG pathway was responsible for this.
will be the subject of our future study. As in other fungi, the A. fumigatus SakA MAPK was transiently phosphorylated by osmotic shock and fungicide treatment, which was fully dependent on SskA. This clearly indicates that under the conditions tested here the SakA MAPK cascade is solely regulated by the TCS. Unexpectedly, we found that SakA was phosphorylated in response to osmotic shock in DnikA cells, suggesting that another mechanism modulates the SakA R428 in vivo phosphorylation instead of NikA, possibly via SskA. On the one hand, NikA seemed to be indispensable for the accumulation of phosphorylated SakA in response to low concentrations of fludioxonil treatment, whereas it was dispensable when treated with 10 mg/ mL fludioxonil. Judged by the roughly estimated growth ICs, 10 mg/mL of fludioxonil seems high and may cause a leaky effect at a specific interaction between the chemical and the target protein. In C. heterostrophus and N. crassa, the accumulation of phosphorylated Hog1-type MAPK was observed in the cells lacking the Nik1-type HK in the presence of high concentrations of fungicides or high osmolar reagents. These results could support the idea that another HK or mechanism contributes to the phosphorylation of SakA via SskA in some filamentous fungi. Notably, in contrast to our results, in a previous study by McCormick et al., TcsC/ NikA has been shown to be required for the phosphorylation of SakA in response to fludioxonil treatment and osmotic shock in A. fumigatus. We think that this discrepancy is derived from the differences in media, cultivation time, and treatment time. Considering these results, the regulation of the SakA MAPK cascade by NikA may be growth stage-specific. We also found that the phosphorylation level of the SakA MAPK in DnikA was moderately high in the absence of stimuli. This increased phosphorylation level is reminiscent of a role of the S. cerevisiae Sln1p in modulating the Hog1 MAPK phosphorylation, where a defect in the sln1 gene causes abnormal activation of the Hog1 MAPK and leads to growth defects. In fact, DnikA showed a slight defect in radial growth on YGMM plates and increased the phosphorylation of SakA. These findings could support the idea that similarly to Sln1p NikA is a negative regulator of the SakA MAPK, whereas it plays a positive role at least in response to fludioxonil treatment. Here, it should be noted that the expression level of catA, dprA, and dprB in DnikA in the absence of stimuli was comparable to that in WT, suggesting that the moderate accumulation of phosphorylated SakA dose not necessarily lead to the high expression of the genes. There may be unknown regulatory mechanisms, for example a negative feedback system, concerning this. In any event, judged by the results of SakA phosphorylation and transcriptional responses to the stimuli, the phosphorylated form of SakA appears to be an active form. Also, we showed a straightforward signaling from SskA to the SakA MAPK in the A. fumigatus HOG pathway both at the transcriptional and post-translational levels. In KRX-0401 hyperosmotic stress conditions, the DnikA, DsskA, and DsakA strains showed growth sensitivity at distinct levels. As has been well studied in S. cerevisiae, the HOG pathway plays an important role in osmotic adaptation by regulating glycerol accumulation; thus, it is possible that glycerol accumulation occurred in response to hyperosmotic stress conditions in A. fumigatus, and the HOG pathway was responsible for this.