As early as 12 hours after axotomy, the dendrites of injured dLGN projection neurons had begun to lose their appendages and distal segments of secondary dendrites. Within the next 48 hours, axotomized projection neurons gradually lost all of the distal segments 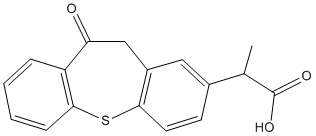 of their dendrites. The sequence of dendritic degeneration, from the loss of appendages and the formation of varicosities to the disappearance of secondary dendrites, generally was consistent among cells in each of the three classes of projection neurons that we have described and from cell to cell within each class. However, the initiation, duration and endpoint of the sequence varied among axotomized dLGN projection neurons. Thus while all dLGN projection neurons in this study experienced a comparable injury and displayed similar structural alterations in their dendrites soon after axotomy, the extent of these alterations varied with the class of projection neuron, and often differed among neurons within each class of projection neurons, regardless of the survival time after axotomy. Among the three classes of projection neurons, cells that retained four or more primary dendrites after axotomy were radial or basket cells. In contrast, bipolar cells experienced severe dendritic degeneration and disappeared from the dLGN earlier than basket or radial cells. These results suggest that the three classes of dLGN projection neurons differ in their vulnerability to axotomy, bipolar cells being more vulnerable than basket cells which in turn are more vulnerable than radial cells. The reasons for this differential vulnerability remain to be determined, but it may be related to the number of afferent synapses that each class of projection neuron can support after axotomy. The number of dendrites, and, therefore, the opportunity for synaptic connections is greater in radial cells than in basket cells and least in bipolar cells. In 3,4,5-Trimethoxyphenylacetic acid summary, dendritic degeneration is a very early event in adult rat dLGN neurons after axotomy. Changes in the dendritic structure of dLGN projection neurons emerge within 12 hours after axotomy. These dendritic changes continue during the next 48 hours, during which time most dLGN projection neurons lose their secondary dendrites. The disappearance of secondary dendrites is accompanied by cell soma atrophy, which is underway 72 hours after axotomy. Since dLGN projection neurons undergo a predictable sequence of structural alterations after axotomy, these alterations may be used as criteria to discriminate among different degenerative stages of injured neurons, and may be useful as reference points to investigate the biochemical and molecular events that underlie cell death after axotomy at defined stages. In this report we also have confirmed caspase-3 activity in axotomized dLGN projection neurons using an antibody raised against an actin fragment, fractin, which results solely from the cleavage of beta actin by activated caspase-3. While calpain may cleave actin in addition to caspase-3, fractin is a specific 32 kD product produced solely by the cleavage of actin by 4-(Benzyloxy)phenol caspase 3 as reported by several previous studies that involved: the immunolabeling of Hirano bodies, which are eosinophilic rodshaped inclusions found primarily in CA1 hippocampal neurons in aged animals and in the brains of Alzheimer��s disease patients; a demonstration that the caspase-specific cleavage product fractin is produced in oligodendroglial cells during TGF-beta-mediated apoptosis; distinguishing between calpain and caspase-3 activity by staining for fractin in excitoxicity.
of their dendrites. The sequence of dendritic degeneration, from the loss of appendages and the formation of varicosities to the disappearance of secondary dendrites, generally was consistent among cells in each of the three classes of projection neurons that we have described and from cell to cell within each class. However, the initiation, duration and endpoint of the sequence varied among axotomized dLGN projection neurons. Thus while all dLGN projection neurons in this study experienced a comparable injury and displayed similar structural alterations in their dendrites soon after axotomy, the extent of these alterations varied with the class of projection neuron, and often differed among neurons within each class of projection neurons, regardless of the survival time after axotomy. Among the three classes of projection neurons, cells that retained four or more primary dendrites after axotomy were radial or basket cells. In contrast, bipolar cells experienced severe dendritic degeneration and disappeared from the dLGN earlier than basket or radial cells. These results suggest that the three classes of dLGN projection neurons differ in their vulnerability to axotomy, bipolar cells being more vulnerable than basket cells which in turn are more vulnerable than radial cells. The reasons for this differential vulnerability remain to be determined, but it may be related to the number of afferent synapses that each class of projection neuron can support after axotomy. The number of dendrites, and, therefore, the opportunity for synaptic connections is greater in radial cells than in basket cells and least in bipolar cells. In 3,4,5-Trimethoxyphenylacetic acid summary, dendritic degeneration is a very early event in adult rat dLGN neurons after axotomy. Changes in the dendritic structure of dLGN projection neurons emerge within 12 hours after axotomy. These dendritic changes continue during the next 48 hours, during which time most dLGN projection neurons lose their secondary dendrites. The disappearance of secondary dendrites is accompanied by cell soma atrophy, which is underway 72 hours after axotomy. Since dLGN projection neurons undergo a predictable sequence of structural alterations after axotomy, these alterations may be used as criteria to discriminate among different degenerative stages of injured neurons, and may be useful as reference points to investigate the biochemical and molecular events that underlie cell death after axotomy at defined stages. In this report we also have confirmed caspase-3 activity in axotomized dLGN projection neurons using an antibody raised against an actin fragment, fractin, which results solely from the cleavage of beta actin by activated caspase-3. While calpain may cleave actin in addition to caspase-3, fractin is a specific 32 kD product produced solely by the cleavage of actin by 4-(Benzyloxy)phenol caspase 3 as reported by several previous studies that involved: the immunolabeling of Hirano bodies, which are eosinophilic rodshaped inclusions found primarily in CA1 hippocampal neurons in aged animals and in the brains of Alzheimer��s disease patients; a demonstration that the caspase-specific cleavage product fractin is produced in oligodendroglial cells during TGF-beta-mediated apoptosis; distinguishing between calpain and caspase-3 activity by staining for fractin in excitoxicity.